Introducción
El agua es indispensable en la vida de las plantas e influye en su crecimiento y productividad. La poca disponibilidad de agua en el suelo puede ocasionar que la transpiración exceda el agua absorbida por las raíces, lo que se conoce como estrés hídrico (Munns y Tester, 2008).
Teniendo en cuenta la gran importancia del agua en las plantas, se puede considerar que una cantidad limitada o excesiva de agua para éstas constituye un factor inductor de situaciones adversas o estresantes (Moreno, 2009).
Por otro lado, una tercera parte de la superficie del planeta se considera como árida o semiárida, mientras que la mayoría de la superficie restante está sujeta a períodos temporales de déficit hídrico. De esta manera, el agua constituye el principal factor limitante del crecimiento de las plantas en la tierra, actuando como una fuerza selectiva de primer grado para la evolución y distribución de las especies vegetales (Hanson y Hitz, 1982).
A lo largo de la evolución, las plantas han desarrollado diferentes respuestas y adaptaciones que les permiten sobrevivir en condiciones de constante déficit hídrico (Nilsen y Orcutt, 1996). Muchas de estas adaptaciones están relacionadas con una mayor capacidad de tomar agua o con un uso más eficiente de este recurso (Moreno, 2009).
Cuando el déficit hídrico se desarrolla lentamente, las plantas pueden presentar respuestas de aclimatación que tienen efectos sobre el crecimiento, como la disminución de la expansión foliar y el aumento del crecimiento radicular (Potters et al., 2007; Shao et al., 2008). Otro mecanismo de resistencia a nivel fisiológico es el cierre de estomas, estructuras responsables de la mayor proporción de pérdida de agua en las plantas (Taiz y Zeiger, 2006).
El siguiente reporte constituye una revisión de algunos artículos científicos y literatura asociada al estrés hídrico en plantas y bosques mediterráneos, sus mecanismos de respuesta y capacidad de adaptación o aclimatación a situaciones adversas de disponibilidad de agua y una breve asociación con los efectos del cambio climático y el bosque esclerófilo chileno.
1. Estrés hídrico y respuesta de las plantas
El estrés por déficit hídrico o por sequía se produce en las plantas en respuesta a un ambiente escaso en agua, en donde la tasa de transpiración excede a la toma de agua. El déficit hídrico no sólo ocurre cuando hay poca agua en el ambiente, sino también por bajas temperaturas y por una elevada salinidad del suelo.
De acuerdo con los requerimientos de agua, las plantas pueden ser consideradas como hidrófitas si están adaptadas a vivir total o parcialmente sumergidas en el agua (en general no toleran potenciales hídricos más negativos de -5 a -10 bares); como mesófitas si están adaptadas a un aporte moderado de agua (en general no toleran potenciales hídricos más negativos de -20 bares) y como xerófitas si están adaptadas a ambientes áridos (en general no toleran potenciales hídricos más negativos de -40 bares) (Nilsen y Orcutt, 1996).
Las plantas a lo largo de su desarrollo experimentan algún grado de estrés por déficit hídrico. En los sistemas naturales, un déficit de agua puede ser el resultado de bajas precipitaciones, baja capacidad de retención de agua del suelo, excesiva salinidad, temperaturas extremas frías o calientes, baja presión de vapor atmosférica o una combinación de estos factores (Nilsen y Orcutt, 1996).
Las plantas han respondido al estrés hídrico desarrollando evolutivamente adaptaciones tanto a nivel morfológico como anatómico y celular, que les permiten vivir en condiciones de constante estrés hídrico (Nilsen y Orcutt, 1996). Las plantas que son capaces de adquirir más agua o que hacen un uso más eficiente de ésta podrán tener resistencia al estrés por sequía.
Las plantas también poseen mecanismos de aclimatación que se activan en respuesta a estrés hídrico (Nilsen y Orcutt, 1996). Cuando el déficit hídrico se desarrolla lentamente, se dan cambios en procesos de desarrollo que tienen varios efectos sobre el crecimiento. Uno de principal importancia es la limitación específica de la expansión foliar. Aunque el área foliar es importante, pues de ella depende la fotosíntesis, una rápida expansión foliar puede afectar negativamente la adaptación a la poca disponibilidad de agua. Otro proceso que se modifica es el crecimiento radicular. La disponibilidad de agua afecta la relación entre el crecimiento de la parte aérea y la raíz; la raíz continúa su desarrollo mientras que la parte aérea deja de crecer por causa del estrés. Así, las plantas son capaces de continuar el desarrollo de sus raíces en búsqueda de agua en zonas más profundas del suelo (Potters et al., 2007; Shao et al., 2008).
Otro mecanismo de resistencia a nivel fisiológico es el cierre de estomas, ya que estos son los responsables de la mayor proporción de pérdida de agua en las plantas (Taiz y Zeiger, 2006).
A nivel celular, otra respuesta de resistencia es el ajuste osmótico, que consiste en una disminución del potencial hídrico en los tejidos vegetales, lo cual tiene como consecuencia la entrada de agua y, por tanto, no se presenta una disminución en el turgor o en la productividad fotosintética.
Una respuesta molecular de las plantas al estrés, y quizá una de las más importantes, es la modificación de la expresión de genes. Durante el déficit hídrico, diferentes tipos celulares responden incrementando o disminuyendo la expresión de algunos genes. Igualmente, se ha visto que muchos genes que no se expresan en condiciones de irrigación óptima pueden empezar a hacerlo bajo déficit hídrico.
2. Transporte de agua en las plantas y potencial hídrico
La planta utiliza la mayor parte del agua (99%) para procesos físicos como disolvente, medio de reacción y a nivel estructural proporcionando turgencia a las estructuras vegetales porque se acumula en vacuolas. Sólo un 1% del agua que absorbe la utiliza como nutriente (Azcón-Bieto, y Talón, 2008). El transporte de agua en la planta se basa en los gradientes de potencial hídrico (Ψ) que se generan en el xilema debido a la transpiración de agua a través de las hojas. El potencial hídrico de define como la capacidad de las moléculas de agua para moverse en un sistema (Azcón-Bieto, y Talón, 2008).
El agua viaja desde las zonas donde el potencial hídrico es mayor (menos negativo) hacia las zonas donde este es menor (más negativo). El potencial hídrico se define según la ecuación: Ψw = p – s donde “Ψw” es el potencial hídrico; “p” es la presión de turgor o la fuerza hidrostática ejercida en la célula vegetal contra la pared celular y es de signo (+), y “s” es la presión osmótica, que es una medida de la concentración de los solutos (Taiz y Zeiger, 2006).
El turgor es, según la ecuación, directamente proporcional al potencial hídrico. La principal fuerza motora que impulsa al agua en su viaje a la parte aérea es la pérdida de agua en las hojas por transpiración. Esto supone que las hojas son los órganos de las plantas que presentan los potenciales hídricos más negativos (Zyalalov, 2004).
La transpiración es la pérdida de agua en forma de vapor por parte de la planta, principalmente a través de las hojas. La transpiración es la responsable de generar un aumento en el gradiente de potencial hídrico en la planta y hace que el agua ascienda desde las raíces (mayor Ψ) a las hojas (menor Ψ) a través del xilema, gracias a las fuerzas de cohesión y tensión características de las moléculas de agua (Fig. 1). El agua se evapora en las hojas a través de los estomas, en mayor o menor medida en función de varios factores (Azcón-Bieto, y Talón, 2008).
El potencial hídrico del suelo, está relacionado con la capacidad de retener agua por parte del suelo y depende del potencial hídrico matricial (Ψm). Cuando las raíces absorben agua del suelo, disminuye su potencial hídrico en las capas superficiales, desplazándose el agua de las zonas más húmedas a las más secas. La velocidad con la que el agua pasa del suelo a las raíces depende del tipo de suelo y de su contenido hídrico. Un suelo arcilloso está formado por partículas más finas que un suelo arenoso, por lo que el primero retiene más agua, siendo su potencial hídrico matricial mayor (más negativo). Por tanto, con la misma cantidad de agua, el potencial hídrico del suelo arcilloso es menor que el arenoso (Azcón-Bieto, y Talón, 2008).
Una vez que el agua ha sido absorbida por las raíces, se moverá por la planta siguiendo dos tipos de transporte pasivo, tratando de alcanzar el equilibrio de potencial hídrico (Fig.1):
- Flujo masivo: movimiento de agua y solutos conjuntamente en una dirección siguiendo las diferencias de presión generadas por la evapotranspiración. Este movimiento es posible gracias a las fuerzas de adhesión y cohesión que establecen las moléculas de agua a través de enlaces de hidrógeno.
- Difusión: responsable del transporte a corta distancia, bien desde el suelo a la planta o entre células.
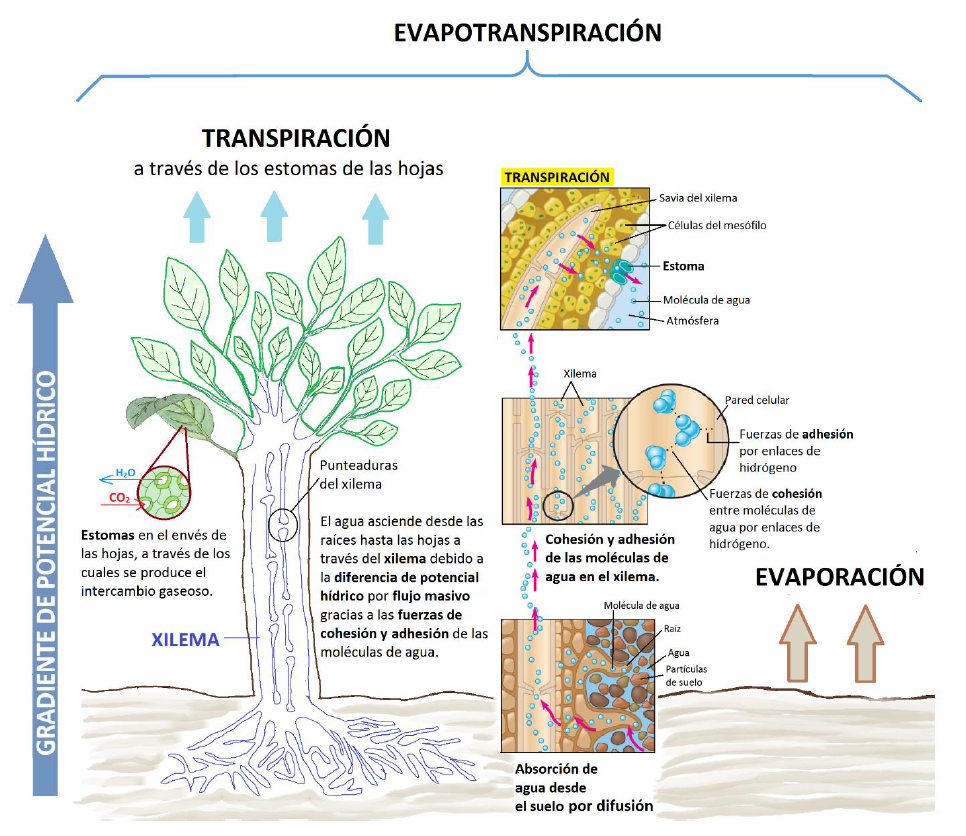
A la izquierda se representa una simplificación de la estructura del xilema, a través del cual el agua asciende desde las raíces hasta las hojas siguiendo un gradiente de potencial hídrico. En este proceso intervendrá tanto el transporte por flujo masivo, que permitirá el ascenso del agua a través del xilema, como la difusión, presente en el transporte de agua entre células y en la absorción de agua por las raíces. A la derecha se representa en detalle el proceso de transporte por flujo masivo, en el que es fundamental las fuerzas de adhesión y cohesión que establecen las moléculas de agua con la pared del xilema y entre ellas respectivamente, gracias a la capacidad de formar enlaces de hidrógeno. Con la apertura estomática tendrá lugar el intercambio gaseoso, liberándose vapor de agua hacia la atmósfera, proceso conocido como transpiración.
Fuente: Carrasco, 2017.Respuesta al estrés hídrico en plantas mediterráneas Perspectiva frente al cambio climático
3. Factores que influyen en la transpiración
De acuerdo a lo señalado por Carrasco (2017) los factores que influyen en la transpiración de las plantas pueden clasificarse en aquellos asociados a factores ambientales, como; la radiación, la presión de vapor del aire, la temperatura, la velocidad del aire y el suministro de agua. Y otros asociados a factores propios de la planta, tales como: la cutícula, los estomas y su área foliar.
A continuación presentamos una síntesis de los principales mecanismos de resistencia de las plantas al estrés hídrico.
3.1. Factores propios de la planta
a) Resistencia cuticular: La cutícula es una capa más o menos gruesa, enriquecida en distinto grado en materiales hidrófobos, de naturaleza cérea, que dificulta la pérdida de agua. Las plantas xerófitas tienen mayor resistencia cuticular que las mesófilas. Si la cutícula actuara como una barrera completa, bloquearía el intercambio gaseoso, esencial para la planta, por lo que esta alcanza un equilibrio utilizando la regulación temporal a través del ajuste estomático (Azcón-Bieto, y Talón, 2008).
b) Regulación estomática: Los estomas son unas aberturas regulables que se encuentran en el tejido epidérmico, principalmente en el envés de la hoja, a través de los que se produce el intercambio gaseoso. El grado de hidratación de la hoja, determina la turgencia de las células alrededor de los estomas (células oclusivas y células epidérmicas adyacentes) y con ello la mayor o menos apertura de los mismos.
Fuente: Azcón-Bieto, y Talón, 2008, citado por Carrasco 2017. En Respuesta al estrés hídrico en plantas mediterráneas Perspectiva frente al cambio climático.
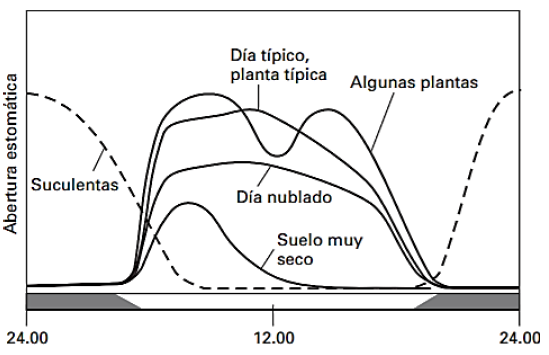
La apertura estomática sigue, normalmente, un ritmo circadiano: los estomas tienen su máxima apertura por la mañana inducida por la luz solar, así se favorece la eficiencia fotosintética de la planta (relación entre la producción energética y la insolación recibida). Además, la evaporación del agua en la superficie de la hoja al producirse la transpiración disminuye su temperatura y así tolera mejor el calor. Los estomas se cerrarán por la noche (Taiz y Zeiger. 2002). Sin embargo, este sistema puede verse alterado en condiciones de déficit hídrico, temperaturas elevadas y/o luz intensa, cuando los estomas pueden cerrarse en las horas centrales del día para evitar la pérdida excesiva de agua. Si el estrés hídrico es aún más acusado, solamente se abrirán a primera hora de la mañana (Fig. 2) ya que durante el día el calor disminuiría la presión de vapor atmosférica, aumentando la transpiración y disminuyendo, por tanto, la eficiencia transpiratoria (pues por cada molécula de CO2 fijado se perdería mucho vapor de agua). Este fenómeno es muy frecuente en la época estival en regiones de clima mediterráneo.
El objetivo del cierre estomático debido al déficit hídrico es aumentar la eficiencia transpiratoria. Sin embargo, el cierre estomático también conlleva una disminución de la absorción de CO2, lo que puede ser contraproducente si nos referimos al aumento de la eficiencia fotosintética que persigue el cierre estomático.
Estas dos formas de resistencia a la desecación tendrán mayor o menor relevancia en función del momento del día: el cierre estomático será más determinante por el día, ya que los estomas se encuentran abiertos para favorecer el proceso de fotosíntesis (captación de CO2); mientras que por la noche, cuando los estomas permanecen cerrados (en plantas C3 y C4), la resistencia cuticular es más determinante para evitar la pérdida de agua (Azcón-Bieto, y Talón, 2008).
c) Reducción del área Foliar: El déficit hídrico producirá una disminución en la turgencia de las células, limitando la expansión. El objetivo es disminuir la tasa transpiratoria y por tanto evitar la pérdida de agua por evaporación (Taiz y Zeiger. 2002).
3.2. Factores ambientales
La presión de vapor (Pv) es la cantidad de agua que contiene el aire; Si la Pv del aire que rodea a la planta es elevada, la transpiración disminuirá y viceversa, ya que se trata de alcanzar un equilibrio entre las presiones. La Pv puede variar, inversamente, en función de la velocidad del aire y de la temperatura. Un aumento de la temperatura disminuirá la Pv y con ello aumentará la transpiración (Taiz y Zeiger. 2002).
Para comprender mejor el efecto de la velocidad del aire sobre la Pv, es necesario definir lo que se conoce como capa límite o capa estacionaria: aire en calma que se encuentra casi saturado de vapor de agua, próximo a la superficie foliar (Taiz y Zeiger. 2002; Azcón-Bieto, y Talón, 2008); la resistencia que ejerce sobre la transpiración, será mayor cuanto más gruesa sea la capa de aire en reposo que está en contacto con la superficie foliar. Por tanto, si la velocidad del aire aumenta, la capa límite será menor aumentando la transpiración.
4. Mecanismos para resistir la sequia
De acuerdo a la literatura consultada las plantas emplearían ciertos mecanismos para resistir al estrés hídrico generado por la sequía o el déficit de agua. Estos mecanismos apuntarían a dos objetivos centrales evitar o tolerar el déficit hídrico.
4.1. Evitación del déficit hídrico en la planta
4.1.1. Escape a la sequía
Es la estrategia que siguen las especies que tratan de completar su ciclo vital en la época húmeda, quedando en forma de latencia en la estación seca. Producen previamente semillas u órganos protegidos. Para ello adelantan la floración evitando la estación seca, como las terófitas. También hay plantas que entran en dormición vegetativa parcial durante el verano (malacófilas xerofíticas deciduas o semidecíduas de verano) o que poseen órganos subterráneos (bulbos y rizomas) como las geófitas.
4.1.2. Conservación del agua
Otras especies siguen estrategias para evitar la pérdida excesiva de agua o para almacenarla.
a) Reducción del área foliar: El déficit hídrico produce un descenso en la turgencia de las células, disminuyendo tanto la expansión foliar como la elongación radicular y por tanto, la tasa de crecimiento. Una menor área foliar transpirará menos, permitiendo conservar más agua. Además, una hoja pequeña minimiza la capa límite con el objetivo de transferir calor de la hoja al aire durante la evaporación. Se debe tener en cuenta que la disminución en la tasa transpiratoria limitará el proceso de fotosíntesis por disminuir la incorporación de CO2.
b) Abscisión foliar: El área foliar de una planta varía tras la maduración de la misma. En situación de déficit hídrico se produce la senescencia e incluso abscisión de las hojas. Al disminuir el número de hojas, lo hará también la superficie de transpiración, lo que supone un ahorro en términos hídricos, para la planta.
c) Cierre estomático: El cierre estomático supone uno de los mecanismos más rápidos para evitar la pérdida de agua en caso de déficit hídrico. Puede producirse por mecanismo hidropasivo: las células oclusivas de los estomas pueden perder agua directamente por evaporación si el aire es muy seco; o por un mecanismo hidroactivo: cuando la planta se encuentra en un estado de deshidratación generalizado, la concentración de solutos aumenta, de forma que sale agua de las células oclusivas hacia otras zonas para equilibrar el potencial osmótico. Este mecanismo hidroactivo, puede ser activado, también, por la acción del ABA ya que origina cambios en la concentración de solutos, como se ha mencionado anteriormente. En ambos casos, la disminución del contenido hídrico de las células oclusivas disminuye su turgencia, produciendo el cierre estomático de forma rápida. Esta regulación estomática está más condicionada por el estado hídrico de la raíz y del suelo que por el de las hojas.
d) Cutícula gruesa: El estrés hídrico induce la producción de una cutícula más gruesa mediante la deposición de ceras sobre la superficie foliar.
e) Disminución de la transpiración: Esto se consigue mediante hojas pequeñas, paralelas a los rayos solares, blanquecinas o de color claro y brillantes para disminuir la temperatura por reflejar la luz. Por otra parte, los aceites esenciales que aumentan la densidad de la capa límite, los estomas en fondo de criptas que mantienen un ambiente húmedo y el enrollamiento de la hoja por pérdida de turgencia, disminuyen la transpiración porque reducen el gradiente de Pv entre la hoja y el aire que la rodea.
f) Almacenamiento de agua: Las plantas suculentas tienen una pérdida de agua por transpiración despreciable debido a la cutícula gruesa que poseen y al cierre estomático por el día (plantas CAM), almacenando el agua en hojas, troncos y raíces muy vacuolados. Además, estas suelen tener raíces superficiales para aprovechar el agua disponible tras las escasas precipitaciones.
4.1.3. Mantenimiento de la absorción de agua
La relación vástago-raíz está regulada por un equilibrio entre la fotosíntesis y la cantidad de agua incorporada. Este equilibrio se altera en situaciones de déficit hídrico, produciéndose un crecimiento radicular hacia zonas húmedas debido a dos motivos: en primer lugar, la inhibición de la expansión foliar disminuye el gasto energético por parte del vástago, de forma que gran parte de los productos generados (foto-asimilados) se redistribuyen hacia las raíces donde favorecen su crecimiento. Este efecto es menos significativo en plantas que se encuentran en periodo reproductivo ya que, en ese caso, dirigen los foto-asimilados hacia los frutos, en mayor medida que hacia las raíces. Esto podría explicar que las plantas sean mucho más sensibles al déficit hídrico durante el periodo reproductivo. En segundo lugar, la disminución del crecimiento radicular hacia zonas secas se debe a una pérdida en la turgencia de esas células, interrumpiendo su expansión y dirigiendo el crecimiento radicular hacia zonas húmedas (Azcón-Bieto, y Talón, 2008).
Las raíces también pueden presentar adaptaciones para evitar el déficit hídrico: las partes de la raíz que atraviesan suelo muy seco pueden estar recubiertas por una capa impermeable para evitar la pérdida de agua. Por otra parte, algunas especies, como las pertenecientes a la familia de las Bromeliaceas, son capaces de aprovechar el agua de la niebla o del rocío gracias a los pelos que poseen en la superficie de sus hojas (Azcón-Bieto, y Talón, 2008).

Fuente: Jardineriaon.com
https://www.jardineriaon.com/la-adaptacion-natural-las-plantas-resistentes-la-sequia.html
4.2. Tolerancia al déficit hídrico
4.2.1. Mantenimiento de la turgencia
Las plantas consiguen mantener la turgencia gracias al ajuste osmótico. La turgencia permite el desarrollo de los procesos habituales de la planta. La disminución en el potencial hídrico que genera la sequía (entre otros factores de estrés) hace que la planta sintetice compuestos osmoprotectores que pueden promover la retención de agua en el citoplasma o estabilizar las membranas. En este ajuste osmótico intervienen iones (K+, Cl-), solutos orgánicos con carga y lo que se conoce como solutos compatibles (moléculas orgánicas como derivados de prolina y betaína). Así mismo, los azúcares aumentarán la viscosidad del líquido intracelular para evitar que colapsen (Azcón-Bieto, y Talón, 2008).
4.2.2. Tolerancia a la desecación
Las plantas se pueden clasificar en dos grupos según el grado de desecación que son capaces de tolerar:
a) Poiquilohídricas: Aquellas cuyos protoplasmas son capaces de soportar deshidrataciones y rehidrataciones sin sufrir daño, bien porque no se vean afectadas o porque sean capaces de recuperarse fácilmente. En este grupo encontramos líquenes, musgos, algas, helechos y algunas plantas vasculares (Azcón-Bieto, y Talón, 2008).
b) Homeohídricas: Lo son la mayoría de las especies de plantas vasculares. No son capaces de soportar potenciales hídricos muy bajos causando en ellas daños irreversibles. Que estas plantas toleren o no la desecación depende, entre otros factores, de la acción del ácido abscísico y de la síntesis o no de “proteínas de deshidratación” (Azcón-Bieto, y Talón, 2008) o dehidrinas. Las dehidrinas son unas proteínas que se expresan en condiciones de déficit hídrico y se encuentran unidas a lípidos de membrana. Pueden estabilizar membranas, proporcionar residuos hidroxilados para unirse a proteínas en lugar del agua y regular el potencial osmótico al actuar como atrapadoras de agua en situaciones de alta osmolaridad (Rangel, 2009).
5. Estrés hídrico y clima mediterráneo
El clima mediterráneo está presente en las zonas que rodean al mar del mismo nombre; también se encuentran regiones de clima mediterráneo en zonas costeras del oeste de América, Australia y África. El clima mediterráneo se caracteriza, a grandes rasgos, por tener periodos con una climatología bastante característica: inviernos fríos, primavera y otoño bastante lluviosos con temperaturas templadas y un verano muy seco que además registra temperaturas bastante elevadas. Estas características que hemos descrito pueden no ser comunes a todas las regiones mediterráneas pues se considera que existe un amplio macrobioclima mediterráneo, cuya característica común es una sequía estival de, al menos, dos meses consecutivos (Carrasco, 2017).
Principalmente es la impredecibilidad y escasez de precipitaciones lo que supone un desafío para las plantas de clima mediterráneo. El objetivo es encontrar un equilibrio entre aprovechar el agua del que dispondrán en un determinado momento a la vez que deben ser capaces de resistir largos periodos de sequía. Estas plantas cuentan con mecanismos de resistencia frente al estrés hídrico, que les permiten controlar la transpiración, pero manteniendo una tasa fotosintética suficiente para vivir.
5. Estrés hídrico y cambio climático
Según Valladares et al. (2004) el clima mediterráneo impone una doble adversidad a los sistemas biológicos: limitación hídrica e irregularidad de las precipitaciones, las que se incrementarán por el cambio climático. De hecho, las cinco regiones mediterráneas del mundo (zonas de Australia, Sudáfrica, Chile, California y la cuenca mediterránea en Europa) se localizan en una franja latitudinal donde la mayor parte de los modelos de circulación atmosférica predicen una disminución de la precipitación durante las próximas décadas (Gracia, 2002; Peñuelas et al., 2004). Además, en estas regiones de alta densidad humana, la biota de los ecosistemas mediterráneos está fuertemente amenazada por la destrucción de los hábitats naturales (Arroyo et al., 2006).
Teniendo en cuenta que el cambio climático tiende a aumentar tanto la temperatura como la irregularidad y escasez de las lluvias, lo que conllevará a un aumento de la duración e intensidad de las sequías, conocer que mecanismos siguen estas plantas frente al estrés hídrico es de suma importancia para ser capaces de gestionar, de la mejor manera posible, los cambios asociados al calentamiento global (Carrasco, 2017).
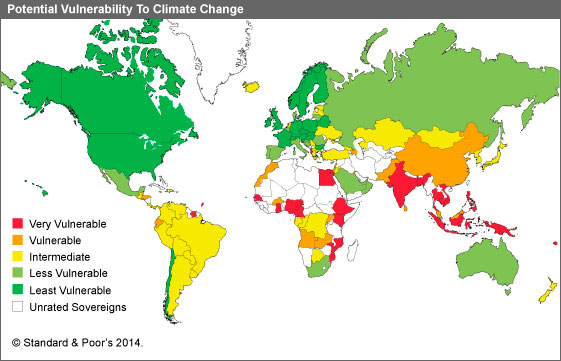
Fuente: The Eco Note, https://thenote.cl/category/conozca-los-paises-mas-vulnerables-al-cambio-climatico/
En el clima mediterráneo, el factor más importante de estrés es el déficit hídrico, el cual se encuentra interrelacionado con otros estreses como temperaturas elevadas, disminución de la humedad o elevada insolación. Con el cambio climático, estos efectos se están acentuando (Valladares et al., 2004), por lo que conocer los mecanismos de resistencia que utilizan las plantas para afrontarlos está cobrando cada vez más interés.
La idea de estrés en general y de estrés hídrico en particular es central para comprender el funcionamiento y la evolución de los sistemas mediterráneos (Mitrakos 1980). A la característica climática general de déficit hídrico, particularmente durante el verano, hay que añadir una notable fluctuación interanual en los regímenes de temperaturas y lluvias, lo cual magnifica el efecto del estrés hídrico al ser poco o nada predecible cuando, con qué intensidad y bajo qué temperaturas tendrá lugar la sequía. Por ejemplo, el porcentaje de variación de las medias móviles ponderadas de 30 años de la pluviometría de la España atlántica es del 11-15%, mientras que es del 25-30% en la España mediterránea (Rodó y Comín 2001). Esta escasa predecibilidad climática hace difícil que los organismos puedan anticiparse al momento más adverso y estar fisiológica y morfológicamente bien preparados. Por ello resulta fácil de justificar el valor adaptativo en zonas de clima mediterráneo de un uso conservativo de los recursos, caracterizado por una respuesta en general poco pronunciada y encaminada más al ahorro y la tolerancia que al gasto y la productividad (Valladares et al. 2000).
El cambio climático consiste en una elevación de la temperatura de la tierra, que parece tener relación con el aumento en la emisión de gases de efecto invernadero. Ante esta situación, el Grupo Intergubernamental de Expertos sobre el Cambio Climático (IPCC), que estudian el impacto del cambio climático en toda su amplitud, han determinado en su quinto informe de evaluación que los ecosistemas de tipo mediterráneo se encuentran entre los más vulnerables debido al incremento de la temperatura, de las sequías y del riesgo de incendios. De hecho, los cambios en las especies dominantes, el aumento de la desertificación, etc. ya son visibles en la actualidad (Camino et al. 2014).
6. Efectos de la sequía y cambio climático en los ecosistemas mediterráneos de Chile.
El bosque esclerófilo chileno se encuentra inserto en un clima de tipo mediterráneo. Durante el periodo estival, la baja disponibilidad hídrica del suelo, en conjunto con las altas temperaturas y radiación solar de la estación, favorecen la aparición del estrés hídrico. Éste es uno de los factores más limitantes del crecimiento, composición y distribución de las especies vegetales, ya que afecta varios aspectos de su funcionamiento, desde el metabolismo celular (incluyendo la fotosíntesis), hasta el crecimiento vegetativo (Cabrera, 2002; Peña-Rojas et al., 2004; Otieno et al., 2005; Flexas et al., 2006, 2014; Fleck et al., 2010; Martínez et al., 2014). El mantenimiento de las funciones vitales de la planta, tales como crecimiento, apertura estomática y fotosíntesis, solo es posible bajo condiciones de estrés hídrico si se mantiene el turgor celular de los tejidos mediante un proceso de ajuste en las propiedades elásticas de la pared celular (ajuste elástico), o la reducción en el potencial osmótico (ajuste osmótico) (Pardos et al., 2005). En este sentido, son escasos los antecedentes respecto a las estrategias que utilizan las especies nativas del bosque mediterráneo chileno para enfrentar períodos de restricción hídrica (Donoso et al., 2011).
No obstante, existen algunos estudios que han evaluado las respuestas de especies del bosque esclerófilo ante situaciones de restricción o estrés hídrico, los cuales revelan algunos de los mecanismos de respuesta que emplean los árboles para mantenerse “vivos”.

Fuente: CR2. Centro de Investigación del Clima y Resiliencia. En: http://www.cr2.cl/la-alarmante-perdida-de-resistencia-del-bosque-esclerofilo-al-menos-un-tercio-ha-disminuido-su-verdor-por-la-megasequia-en-la-zona-central-ladera-sur/
Por ejemplo, Donoso et al (2011) evaluaron la respuesta fisiológica y de crecimiento en plantas de Quillaja saponaria (quillay) y Cryptocarya alba (peumo) sometidas a restricción hídrica, obteniendo como resultados que quillay, al enfrentarse al periodo de estrés más intenso, disminuye su biomasa foliar para equilibrar las pérdidas por transpiración, con la eficiencia del sistema radicular de suministrar agua a las hojas. Peumo en tanto, presenta una tendencia hacia el ajuste osmótico para mantener altos valores de contenido hídrico relativo y turgencia celular. Así mismo, en su evaluación los autores detectaron que la fotosíntesis neta, conductancia estomática y transpiración, experimentaron una drástica reducción en sus valores en las plantas bajo riego restringido respecto a las plantas del tratamiento testigo. En consecuencia, las plantas con riego restringido presentan un crecimiento y acumulación de biomasa sustantivamente menor al que presentan las plantas con riego constante. Bajo condiciones de déficit hídrico, quillay disminuye significativamente la relación biomasa aérea/biomasa radical, debido a la defoliación que sufre y al incremento de la biomasa de raíces, mientras que en peumo la relación permanece invariable.

Fuente: CR2. Centro de Investigación del Clima y Resiliencia. En: http://www.cr2.cl/la-alarmante-perdida-de-resistencia-del-bosque-esclerofilo-al-menos-un-tercio-ha-disminuido-su-verdor-por-la-megasequia-en-la-zona-central-ladera-sur/
Por otro lado, Peña-Rojas et al (2017) investigaron las respuestas de Peumus boldus (boldo) obteniendo como resultado que la especie mantendría un balance de carbono positivo, aún bajo condiciones de estrés, lo que permitiría mantener la función de crecimiento, aunque a una tasa reducida. Esto implica una mayor tolerancia a períodos de estrés hídrico, y permitiría a la especie ocupar zonas con menor disponibilidad hídrica, explicando su dominancia en algunos sitios dentro de la asociación Cryptocarya alba – Lithraea caustica (peumo – litre) y, por otra parte, su aparición aislada en laderas más secas junto a Quillaja saponaria (quillay), Lithrea caustica (litre), Acacia caven (espino) y Retanilla trinervia (Tevo) (Donoso, 1982).
7. Conclusiones
- A lo largo de la evolución, las plantas han desarrollado diferentes respuestas y adaptaciones que les permiten sobrevivir en condiciones de constante déficit hídrico. Muchas de estas adaptaciones están relacionadas con una mayor capacidad de tomar agua o con un uso más eficiente de este recurso.
- El estrés por déficit hídrico o por sequía se produce en las plantas en respuesta a un ambiente escaso en agua, en donde la tasa de transpiración excede a la toma de agua. El déficit hídrico no sólo ocurre cuando hay poca agua en el ambiente, sino también por bajas temperaturas y por una elevada salinidad del suelo.
- Cuando el déficit hídrico se desarrolla lentamente, las plantas pueden presentar respuestas de aclimatación que tienen efectos sobre el crecimiento, como la disminución de la expansión foliar y el aumento del crecimiento radicular.
- La planta utiliza la mayor parte del agua (99%) para procesos físicos como disolvente, medio de reacción y a nivel estructural proporcionando turgencia a las estructuras vegetales porque se acumula en vacuolas. Sólo un 1% del agua que absorbe la utiliza como nutriente.
- El transporte de agua en la planta se basa en los gradientes de potencial hídrico (Ψ) que se generan en el xilema debido a la transpiración de agua a través de las hojas. El potencial hídrico de define como la capacidad de las moléculas de agua para moverse en un sistema.
- Los factores que influyen en la transpiración de las plantas pueden clasificarse en aquellos asociados a factores ambientales, como; la radiación, la presión de vapor del aire, la temperatura, la velocidad del aire y el suministro de agua. Y otros asociados a factores propios de la planta, tales como: la cutícula, las estomas y su área foliar.
- Las plantas emplean ciertos mecanismos para resistir al estrés hídrico generado por la sequía o el déficit de agua. Estos mecanismos apuntarían a dos objetivos centrales evitar o tolerar el déficit hídrico.
- Dentro de los mecanismos para evitar el déficit hídrico se encuentran aquellos que buscan “escapar” de la sequía completando su ciclo de vida en la época húmeda. Otros que apuntan “conservar” el agua mediante la reducción del área foliar, la abscisión o caída de hojas o el cierre estomático, entre otras. Y los últimos que apuntan a “mantener” la absorción del agua mediante la búsqueda de un equilibrio entre el crecimiento foliar y radicular. Para tolerar el déficit hídrico las plantas buscarán mantener su turgencia gracias a un ajuste osmótico o algunas han logrado adaptarse a para resistir la desecación.
- El clima mediterráneo impone una doble adversidad a los sistemas biológicos: limitación hídrica e irregularidad de las precipitaciones, las que se incrementarán por el cambio climático.
- El cambio climático tiende a aumentar tanto la temperatura como la irregularidad y escasez de las lluvias, lo que conllevará a un aumento de la duración e intensidad de las sequías.
- En el clima mediterráneo, el factor más importante de estrés es el déficit hídrico, el cual se encuentra interrelacionado con otros estreses como temperaturas elevadas, disminución de la humedad o elevada insolación.
- Existen algunos estudios que han evaluado las respuestas de especies del bosque esclerófilo chileno ante situaciones de restricción o estrés hídrico, los cuales revelan algunos de los mecanismos de respuesta que emplean los árboles para mantenerse “vivos” y resistir los períodos de sequía.
8. Bibliografía
Arroyo M, Marquet P, Marticorena C, Simonetti J, Cavieres L, Squeo F, Rozzi R, Massardo F (2006) El Hotspot chileno, prioridad mundial para la conservación. En Saball P, Arroyo M, Castilla JC, Estades C, De Guevara JM, Larraín S, Moreno C, Rivas F, Rovira J, Sánchez A, Sierralta L (Eds.) Biodiversidad de Chile. Patrimonio y Desafíos. Comisión Nacional del Medio Ambiente. Santiago, Chile. pp. 90-93.
Azcón-Bieto, J. y Talón M. 2008. Fundamentos de fisiología vegetal 2ªEd. McGraw-Hill Interamericana de España, S.A.U., Madrid.
Cabrera H (2002) Respuestas ecofisiológicas de plantas en ecosistemas de zonas con clima mediterráneo y ambientes de alta montaña. Rev. Chil. Hist. Nat. 75: 625-637.
Camino, E.R., Ruggeroni, J.R.P. y Hernández, F.H. 2014. Quinto informe de evaluación del IPCC: Bases físicas. Tiempo y Clima, 11(1)
Carrasco, J. 2017. Respuesta al estrés hídrico en plantas mediterráneas Perspectiva frente al cambio climático. Trabajo fin de Grado. Facultad de farmacia, Universidad Complutense.
Donoso C (1982) Reseña ecológica de los bosques mediterráneos de Chile. Bosque 4: 117-146.
Donoso, Sergio, Peña, Karen, Pacheco, Cristian, Luna, Gabriela, & Aguirre, Aldo. (2011). Respuesta fisiológica y de crecimiento en plantas de Quillaja saponaria y Cryptocarya alba sometidas a restricción hídrica. Bosque (Valdivia), 32(2), 187-195. https://dx.doi.org/10.4067/S0717-92002011000200009
Fleck I, Peña-Rojas K, Aranda X (2010) Mesophyll conductance to CO2 and leaf morphological characteristics under drought stress during Quercus ilex L. resprouting. Ann. Forest Sci. 67: 308.
Flexas J, Bota J, Galmés J, Medrano H, Ribas-Carbó M (2006) Keeping a positive carbon balance under adverse conditions: responses of photosynthesis and respiration to water stress. Physiol. Plant. 127: 343-352.
Gracia CA, Sabaté S, Sánchez A (2002) El cambio climático y la reducción de la reserva de agua en el bosque mediterráneo. Ecosistemas 11(2) www.aeet.org/ecosistemas/022/ investigacion4.htm
Karen Peña-Rojas, Sergio Donoso, Rodrigo Gangas, Sergio Durán y David Ilabaca. (2017). EFECTOS DE LA SEQUÍA EN LAS RELACIONES HÍDRICAS, CRECIMIENTO Y DISTRIBUCIÓN DE BIOMASA EN PLANTAS DE Peumus boldus MOLINA (MONIMIACEAE) CULTIVADAS EN VIVERO. http://repositorio.uchile.cl/bitstream/handle/2250/150443/Efectos-de-la-sequia.pdf?sequence=1&isAllowed=y
Martínez C, Cantón M, Juñent F (2014) Incidencia del déficit hídrico en el crecimiento de árboles de uso urbano en ciudades de zonas áridas. Caso de Mendoza, Argentina. Interciencia 39: 890-897.
Moreno, Liz Patricia. 2009. Respuesta de las plantas al estrés por déficit hídrico. Una revisión. Plant responses to water deficit stress. A review. Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. Agronomía Colombiana 27(2), 179-191, 2009.
Mitrakos, K. 1980. A theory for Mediterranean plant life. Acta Oecologica 1: 245-252.
Munns, R. and M. Tester. 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 59: 651-681.
Nilsen, E.T. y D.M. Orcutt. 1996. Physiology of plants under stress. Abiotic factors. John Wiley and Sons, New York, NY.
Otieno DO, Schmidt MW, Adiku S, Tenhunen J (2005) Physiological and morphological responses to water stress in two Acacia species from contrasting habitats. Tree Physiol. 25: 361-371
Pardos M, Jiménez MD, Aranda I, Puértolas J, Pardos JA (2005) Water relations of cork oak (Quercus suber L.) seedlings in response to shading and moderate drought. Ann. Forest Sci. 62: 377-384.
Peña-Rojas K, Aranda X, Fleck I (2004) Stomatal limitation to CO2 assimilation and down-regulation of photosynthesis in Quercus ilex resprouts in response to slowly imposed drought. Tree Physiol. 24: 813-822.
Potters, G., T.P. Pasternak, Y. Guisez, K.J. Palme y M.A.K. Jansen. 2007. Stress-induced morphogenic responses: growing out of trouble? Trends Plant Sci. 12(3), 99-105.
Rangel Fajardo, M.A. 2009. Anatomía y tolerancia a la desecación de las semillas de cacao. Tesis Doctoral, Colegio de postgraduados Montecillo, México.
Rivas-Martínez, S. 2000. Worldwide Bioclimatic Classification System. Phytosociological Research Centre, Universidad Complutense de Madrid, Spain.
Shao, H.B., L.Y. Chu, C.A. Jaleel y C.X. Zhao. 2008. Water-deficit stress-induced anatomical changes in higher plants. C.R. Biol. 331, 215-225.
Taiz, L. and E. Zeiger. 2002. Plant physiology. Sinauer Associates. Sunderland, MA, USA.
Valladares, F., E. Martinez-Ferri, L. Balaguer, E. Perez-Corona y E. Manrique. 2000. Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: a conservative resource-use strategy? New Phytologist 148: 79-91.
Valladares F, Vilagrosa A, Peñuelas J, Ogaya R, Camarero J, Corcuera L, Sisó S, Gil-Pelegrín E (2004) Estrés hídrico: ecofisiología y escalas de sequía. En Valladares F (Eds.) Ecología del Bosque Mediterráneo en un Mundo Cambiante. Ministerio de Medio Ambiente. EGRAF. Madrid, España. pp. 163-190.
Visítanos en nuestras redes